Product Sheet CP10334
Description
BACKGROUND Histones are chromatin proteins that play an important role in DNA packaging and gene regulation. Histones are subjected to a wide variety of covalent modifications that include acetylation, phosphorylation and methylation. Distinct combinatorial patterns of such modifications are believed to serve as epigenetic marks that control chromatin packaging and subsequent gene expression by providing recognition sites for downstream chromatin regulatory factors. Histone methylation plays an important role in many biological processes including heterochromatin formation, homeotic gene silencing, X-chromosome inactivation, genomic imprinting and transcriptional regulation and may exist on both the lysine (K) and arginine (R) residues of histones. Lysine methylation can occur on a variety of specific sites on histone H3 (K4, K9, K27, K36, and K79) and histone H4 (K20), thus allowing for the generation of distinct histone methylation patterns that directly influence chromatin regulated cellular processes. Importantly, lysine residues can also be mono-, di-, or trimethylated, and such differential methylation states serve to diversify the docking sites for effector chromatin proteins and modifiers, thus underscoring the complexity of histone methylation in regulating biological processes. Histone methylation had long been considered an irreversible epigenetic mark, however this viewpoint was challenged with the discovery of the first H3-K4 and K9 specific histone demethylase (HDM) LSD1. Since then, numerous different HDMs have been discovered that display distinct substrate and methylation conversion state specificity, supporting the concept that histone methylation, like acetylation, is a reversible and dynamically regulated process. Investigation of the specific biological roles of these newly identified HDMs will undoubtedly contribute to our understanding of histone methylation regulated cellular processes in development and disease.1
The JmjC-domain-containing histone demthylases (JHDMs) represent the largest class of HDMs. These HDM enzymes are characterized by containing a conserved JmjC domain that catalyzes lysine demethylation of histones via an oxidative reduction reaction that requires iron Fe(II) and alpha-ketoglutarate (aKG) cofactors. Unlike LSD1, that reduces only mono- and dimethyl lysine modifications, certain JHDM family members can also reduce tri- histone lysine-methylation states. Additionally, different JHDM families display distinct substrate specificity. For instance, the JHDM1 family reduces H3K36, the JHDM2 family reduces H3K9 while certain members of the JHDM3/JMJD2 family can reduce both H3K9 and H3K36.2 There are four JHDM3/JMJD2 genes within the human genome, designated JHDM3/JMJD2A-D (here, they will be referred to as JMJD2A-D for simplicity). It has been suggested that JMJD2D gave rise to two additional human genes, JMJD2E and JMJD2F via local retrotransposition. Genes JMJD2A-C encode for proteins containing N-terminal JmjC and JmjN domains, followed by two C-terminus PhD domains and two Tudor domains. In contrast, the JMJD2D family member encodes for a shorter protein product that lacks the C-terminal PHD and Tudor domains. Prior in vitro analysis of the catalytic activity of the four human JMJD2A-D protein family members reveals differences in both their substrate specificity and their ability to mediate different degrees of demethylation, supporting distinct biological functions for these family members. For example, while all JMJD2 family members can reduce H3-K9Me1, only JMJD2A and C have the capacity to also reduce H3-K36Me3. Additionally, while JMJD2A-D can all convert H3-K9Me3 to H3-K9Me2, only JMJD2D can reduce H3-K9Me3 to H3-K9Me2 and H3-K9Me1.3
The JmjC-domain-containing histone demthylases (JHDMs) represent the largest class of HDMs. These HDM enzymes are characterized by containing a conserved JmjC domain that catalyzes lysine demethylation of histones via an oxidative reduction reaction that requires iron Fe(II) and alpha-ketoglutarate (aKG) cofactors. Unlike LSD1, that reduces only mono- and dimethyl lysine modifications, certain JHDM family members can also reduce tri- histone lysine-methylation states. Additionally, different JHDM families display distinct substrate specificity. For instance, the JHDM1 family reduces H3K36, the JHDM2 family reduces H3K9 while certain members of the JHDM3/JMJD2 family can reduce both H3K9 and H3K36.2 There are four JHDM3/JMJD2 genes within the human genome, designated JHDM3/JMJD2A-D (here, they will be referred to as JMJD2A-D for simplicity). It has been suggested that JMJD2D gave rise to two additional human genes, JMJD2E and JMJD2F via local retrotransposition. Genes JMJD2A-C encode for proteins containing N-terminal JmjC and JmjN domains, followed by two C-terminus PhD domains and two Tudor domains. In contrast, the JMJD2D family member encodes for a shorter protein product that lacks the C-terminal PHD and Tudor domains. Prior in vitro analysis of the catalytic activity of the four human JMJD2A-D protein family members reveals differences in both their substrate specificity and their ability to mediate different degrees of demethylation, supporting distinct biological functions for these family members. For example, while all JMJD2 family members can reduce H3-K9Me1, only JMJD2A and C have the capacity to also reduce H3-K36Me3. Additionally, while JMJD2A-D can all convert H3-K9Me3 to H3-K9Me2, only JMJD2D can reduce H3-K9Me3 to H3-K9Me2 and H3-K9Me1.3
REFERENCES
1. Daniel, J.A. et al: Cell Cycle 4:919-26, 2005
2. Huang, Y. et al: Science 312:748-51, 2006
3. Marmorstein, R. & Trievel, R.C.: Biochim. Biophys. Acta 1789:58-68, 2009
2. Huang, Y. et al: Science 312:748-51, 2006
3. Marmorstein, R. & Trievel, R.C.: Biochim. Biophys. Acta 1789:58-68, 2009
Products are for research use only. They are not intended for human, animal, or diagnostic applications.
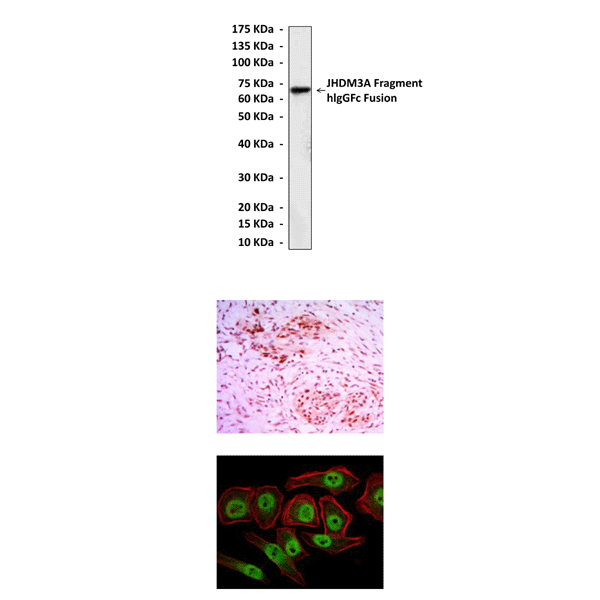
(Click to Enlarge) Top: Western Blot detection of JHDM3A fragment-hIgGFc fusion proteins in 293 cell lysate containing JHDM3A fragment-hIgGFc fusion proteins using JHDM3A Antibody. Middle: This antibody stains paraffin-embedded human colon cancer tissue in immunohistochemical analysis. Bottom: It also stains NTERA-2 cells in confocal immunofluorescent analysis (JHDM3A Antibody: Green; Actin filaments: Red; DRAQ5 DNA Dye: Blue).
Details
Cat.No.: | CP10334 |
Antigen: | Raised against recombinant human JHDM3A fragments expressed in E. coli. |
Isotype: | Mouse IgG1 |
Species & predicted species cross- reactivity ( ): | Human, Mouse, Rat |
Applications & Suggested starting dilutions:* | WB 1:1000 IP n/d IHC 1:50 - 1:200 ICC 1:50 - 1:200 FACS n/d |
Predicted Molecular Weight of protein: | 145 kDa |
Specificity/Sensitivity: | Detects endogenous JHDM3A proteins without cross-reactivity with other family members. |
Storage: | Store at -20°C, 4°C for frequent use. Avoid repeated freeze-thaw cycles. |
*Optimal working dilutions must be determined by end user.
Products
| Product | Size | CAT.# | Price | Quantity |
|---|---|---|---|---|
| Mouse JHDM3A/JMJD2A/KDM4A Antibody: Mouse JHDM3A/JMJD2A/KDM4A Antibody | Size: 100 ul | CAT.#: CP10334 | Price: $413.00 |