Product Sheet CP10344
Description
BACKGROUND Sox (SRY-related HMG box) proteins are a group of transcriptional regulators having analogous HMG domains for their DNA binding, and are highly involved in regulation of cell differentiation. There are roughly 20 Sox genes in each vertebrate species, which are classified into nine groups, according to the organization of the encoding proteins and the HMG domain sequences. Cumulative evidence indicates that Sox proteins form a complex with their partner transcription factors unique to each Sox group, thereby selecting group-specific and partner-dependent regulatory target genes.1
Sox2 belongs to the Sox B1 subgroup, which also includes Sox1 and Sox3, based on homology within and outside the HMG box. Several lines of evidence indicate that Sox2 may act to maintain or preserve developmental potential. Its function is to control pluripotency, in both embryonic stem cell (ESC) and adult-tissue specific stem cell (SC), such as neural SC. Likewise, Sox2 is one of the “Yamanaka” factors and can cooperate with c-Myc, Oct4 and KLF4 to reprogram adult somatic cells into an induced pluripotent state.2 It was also shown that Sox2 marks the pluripotent lineage of the early mouse embryo, so that like Oct4 it is expressed in the inner cell mass (ICM), epiblast, and germ cells. Unlike Oct4, however, Sox2 is also expressed by the multipotential cells of the extraembryonic ectoderm (ExE). In both lineages its down-regulation correlates with a commitment to differentiate, such that it is no longer expressed in cell types with restricted developmental potential. Moreover, it was demonstrated that Oct-4 is a member of the gene family regulated by Oct-4 and Sox2, as reported before for the FGF-4, UTF1, Sox2, and Fbx15 genes. Thus, Oct-4 and Sox2 comprise a regulatory complex that controls the expression of genes important for the maintenance of the primitive state, including themselves. This autoregulatory circuit of the Sox2•Oct-4 complex may contribute to maintaining robustly the precise expression level of Oct-4 in primitive cells.3
Among the Sox genes, group B1 are expressed widely in the nervous system, and are implicated in neural development. In particular, Sox2 expression consistently marks neural primordial cells at various stages of development. For example, Sox2 expression is associated with uncommitted dividing stem and precursor cells of the developing central nervous system (CNS) and indeed can be used to isolate such cells. Furthermore, a recent report investigating the neural primordia of embryos indicates that Sox2 expression is highly correlated with the multipotent neural stem cell state. Because Sox2 is expressed more or less uniformly in the early neural tube, it is regarded as a “pan-neural” marker in early embryonic stages. In addition, Sox2 is commonly expressed in sensory placodes. Another important aspect of Sox2 regulation is that its expression in the CNS is first activated upon neural induction elicited by signals from the organizer. Therefore, initiation of Sox2 expression must be an essential part of the mechanism of neural induction.4 These aspects of Sox2 expression in the early embryo are common among vertebrates. In addition, Sox2 is crucial for morphogenesis and homeostasis of the esophageal, tracheo-bronchial and bronchiolar epithelia. Moreover, several studies reported the identification of Sox2 as a novel master oncogene in lung and esophageal squamous cell carcinoma (SCC) using a screening strategy based on tumor genome analyses.
Sox2 belongs to the Sox B1 subgroup, which also includes Sox1 and Sox3, based on homology within and outside the HMG box. Several lines of evidence indicate that Sox2 may act to maintain or preserve developmental potential. Its function is to control pluripotency, in both embryonic stem cell (ESC) and adult-tissue specific stem cell (SC), such as neural SC. Likewise, Sox2 is one of the “Yamanaka” factors and can cooperate with c-Myc, Oct4 and KLF4 to reprogram adult somatic cells into an induced pluripotent state.2 It was also shown that Sox2 marks the pluripotent lineage of the early mouse embryo, so that like Oct4 it is expressed in the inner cell mass (ICM), epiblast, and germ cells. Unlike Oct4, however, Sox2 is also expressed by the multipotential cells of the extraembryonic ectoderm (ExE). In both lineages its down-regulation correlates with a commitment to differentiate, such that it is no longer expressed in cell types with restricted developmental potential. Moreover, it was demonstrated that Oct-4 is a member of the gene family regulated by Oct-4 and Sox2, as reported before for the FGF-4, UTF1, Sox2, and Fbx15 genes. Thus, Oct-4 and Sox2 comprise a regulatory complex that controls the expression of genes important for the maintenance of the primitive state, including themselves. This autoregulatory circuit of the Sox2•Oct-4 complex may contribute to maintaining robustly the precise expression level of Oct-4 in primitive cells.3
Among the Sox genes, group B1 are expressed widely in the nervous system, and are implicated in neural development. In particular, Sox2 expression consistently marks neural primordial cells at various stages of development. For example, Sox2 expression is associated with uncommitted dividing stem and precursor cells of the developing central nervous system (CNS) and indeed can be used to isolate such cells. Furthermore, a recent report investigating the neural primordia of embryos indicates that Sox2 expression is highly correlated with the multipotent neural stem cell state. Because Sox2 is expressed more or less uniformly in the early neural tube, it is regarded as a “pan-neural” marker in early embryonic stages. In addition, Sox2 is commonly expressed in sensory placodes. Another important aspect of Sox2 regulation is that its expression in the CNS is first activated upon neural induction elicited by signals from the organizer. Therefore, initiation of Sox2 expression must be an essential part of the mechanism of neural induction.4 These aspects of Sox2 expression in the early embryo are common among vertebrates. In addition, Sox2 is crucial for morphogenesis and homeostasis of the esophageal, tracheo-bronchial and bronchiolar epithelia. Moreover, several studies reported the identification of Sox2 as a novel master oncogene in lung and esophageal squamous cell carcinoma (SCC) using a screening strategy based on tumor genome analyses.
REFERENCES
1. Episkopou, V.: Trends Neurosci. 28:219-21, 2005
2. Takahashi, K. et al: Cell 131:861-72, 2007
3. Okumura-Nakanishi, S. et al: J. Biol. Chem. 280:5307-17, 2005
4. Uchikawa, M. et al: Dev. Cell 4:509-19, 2003
5. Hussenet, T. & du Manoir, S.: Cell Cycle 9:1480-6, 2010
2. Takahashi, K. et al: Cell 131:861-72, 2007
3. Okumura-Nakanishi, S. et al: J. Biol. Chem. 280:5307-17, 2005
4. Uchikawa, M. et al: Dev. Cell 4:509-19, 2003
5. Hussenet, T. & du Manoir, S.: Cell Cycle 9:1480-6, 2010
Products are for research use only. They are not intended for human, animal, or diagnostic applications.
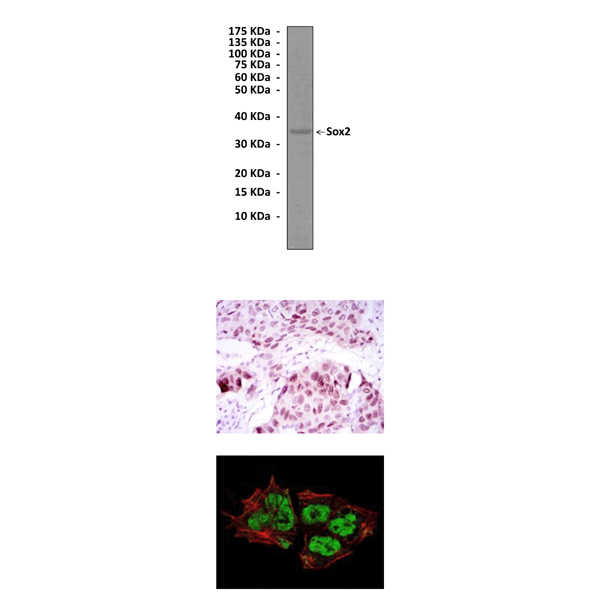
(Click to Enlarge) Top: Western Blot detection of Sox2 proteins in NTERA2 cell lysate using Sox2 Antibody. Middle: Immunohistochemical analysis of paraffin-embedded lung cancer tissues using Sox2 Antibody. Bottom: This antibody also stains NTERA-2 cells in immunofluorescence analysis (Sox2 Antibody: green; Actin filaments: red).
Details
Cat.No.: | CP10344 |
Antigen: | Raised against recombinant human Sox2 fragments expressed in E. coli. |
Isotype: | Mouse IgG1 |
Species & predicted species cross- reactivity ( ): | Human, Mouse, Rat |
Applications & Suggested starting dilutions:* | WB 1:1000 IP n/d IHC n/d ICC n/d FACS n/d |
Predicted Molecular Weight of protein: | 35 kDa |
Specificity/Sensitivity: | detects endogenous Sox2 proteins without cross-reactivity with other family members. |
Storage: | Store at -20°C, 4°C for frequent use. Avoid repeated freeze-thaw cycles. |
*Optimal working dilutions must be determined by end user.
Products
| Product | Size | CAT.# | Price | Quantity |
|---|---|---|---|---|
| Mouse Sox2 Antibody: Mouse Sox2 Antibody | Size: 100 ul | CAT.#: CP10344 | Price: $354.00 |