Product Sheet CP10179
Description
BACKGROUND The MYST proteins, characterized by a highly conserved acetyltransferase domain, are involved in a wide range of physiological processes in mammals, ranging from the development of the nervous system to that of the hematopoietic system. Mof (males absent on the first; also called MYST1 or KAT8) is one of the five mammalian MYST family histone acetyltransferases. Mof has an amino-terminal chromodomain, reported to bind noncoding RNA, and a central MYST histone acetyltransferase domain. This domain structure is identical to that of Tip60 (human immunodeficiency virus Tat-interacting protein 60), and together, Mof and Tip60 form a subclass of MYST histone acetyltransferases.1
In mammalian cell culture systems, a variety of different activities have been ascribed to Mof, including roles in cell cycle regulation and response to DNA repair. Mof can acetylate ATM, a protein with a key role in cell cycle checkpoint control, and through acetylation, modify its activity. Moreover, Mof can acetylate p53 at lysine 120, and this activity is important in directing the cell into an apoptotic pathway via the induction of Bax and Puma gene expression by p53, at least in H1299 cells. Another function ascribed to Mof is to enhance transcription as a coactivator for the trithorax group protein MLL in inducing the expression of the Hoxa9 gene. This action at a specific locus would mean that Mof may have distinctly different molecular roles in mammalian cells from those in insect cells, namely, effects on specific loci versus large regions of the genome, such as the male X chromosome.2
In mammals, the Mof gene is ubiquitously expressed, and most tissues have similar, modest levels of expression. Exceptionally high levels of expression are found in the testis. Mof is expressed in both proliferating and postmitotic cells and, during development, does not appear to be restricted to regions with high levels of apoptosis or restricted to cells progressing through the cell cycle. The wide range of cellular processes that were reported to be affected by Mof, together with ubiquitous expression of the Mof gene, suggests that Mof is a multifunctional protein. A major biochemical role of Mof in Drosophila and mammalian cells is histone 4 lysine 16 (H4K16) acetylation. It was shown that Mof mutant embryos first lack acetylation specifically on H4K16 and then show abnormal chromatin morphology before finally undergoing death by apoptosis.3
In mammalian cell culture systems, a variety of different activities have been ascribed to Mof, including roles in cell cycle regulation and response to DNA repair. Mof can acetylate ATM, a protein with a key role in cell cycle checkpoint control, and through acetylation, modify its activity. Moreover, Mof can acetylate p53 at lysine 120, and this activity is important in directing the cell into an apoptotic pathway via the induction of Bax and Puma gene expression by p53, at least in H1299 cells. Another function ascribed to Mof is to enhance transcription as a coactivator for the trithorax group protein MLL in inducing the expression of the Hoxa9 gene. This action at a specific locus would mean that Mof may have distinctly different molecular roles in mammalian cells from those in insect cells, namely, effects on specific loci versus large regions of the genome, such as the male X chromosome.2
In mammals, the Mof gene is ubiquitously expressed, and most tissues have similar, modest levels of expression. Exceptionally high levels of expression are found in the testis. Mof is expressed in both proliferating and postmitotic cells and, during development, does not appear to be restricted to regions with high levels of apoptosis or restricted to cells progressing through the cell cycle. The wide range of cellular processes that were reported to be affected by Mof, together with ubiquitous expression of the Mof gene, suggests that Mof is a multifunctional protein. A major biochemical role of Mof in Drosophila and mammalian cells is histone 4 lysine 16 (H4K16) acetylation. It was shown that Mof mutant embryos first lack acetylation specifically on H4K16 and then show abnormal chromatin morphology before finally undergoing death by apoptosis.3
REFERENCES
1. Avvakumov, N. & Cote, J. : Oncogene 26:5395-5407, 2007
2. Rea, S. et al: Oncogene 26:5385-94, 2007
3. Yang, X.J.: Neucleic Acid Res. 32:959-76, 2004
2. Rea, S. et al: Oncogene 26:5385-94, 2007
3. Yang, X.J.: Neucleic Acid Res. 32:959-76, 2004
Products are for research use only. They are not intended for human, animal, or diagnostic applications.
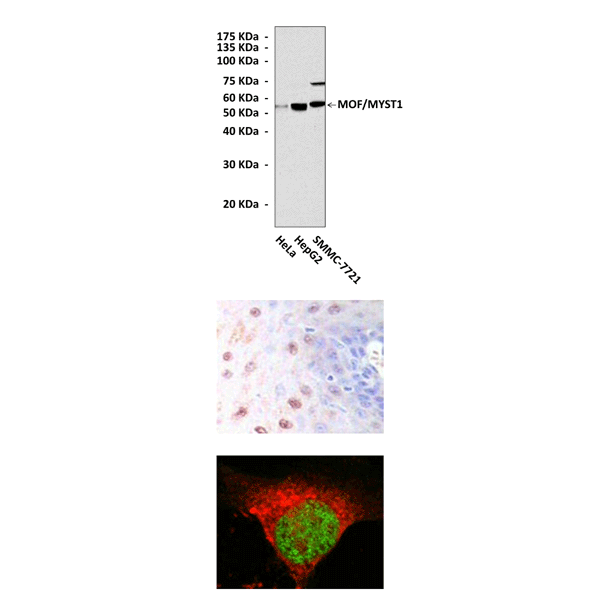
Top: Western Blot detection of MOF proteins in various cell lysates using MOF Antibody. Middle: This antibody stains paraffin-embedded human rectum cancer tissue in immunohistochemical analysis. Bottom: It also stains Eca109 cells in confocal immunofluorescent testing (MOF Antibody: Green; Actin filaments: Red).
Details
Cat.No.: | CP10179 |
Antigen: | Purified recombinant human MOF fragments expressed in E. coli. |
Isotype: | Mouse IgG2b |
Species & predicted species cross- reactivity ( ): | Human |
Applications & Suggested starting dilutions:* | WB 1:1000 IP n/d IHC 1:200 ICC 1:200 FACS n/d |
Predicted Molecular Weight of protein: | 53 kDa |
Specificity/Sensitivity: | Detects endogenous MOF proteins without cross-reactivity with other family members. |
Storage: | Store at -20°C, 4°C for frequent use. Avoid repeated freeze-thaw cycles. |
*Optimal working dilutions must be determined by end user.
Products
| Product | Size | CAT.# | Price | Quantity |
|---|---|---|---|---|
| Mouse MOF/MYST1 Antibody: Mouse MOF/MYST1 Antibody | Size: 100 ul | CAT.#: CP10179 | Price: $413.00 |